Možnosti uplatňování genetiky v holubářské praxi II
Pokračování Cílem druhé části příspěvku je uvést několik praktických příkladů z genetiky zbarvení peří u holubů a vysvětlit metodu používání zápisu genetických vzorců pro popsání základních barev a jednotlivých kreseb či vzorků na opeření holubů. Barva peří na holubech je dána jednak barvou pigmentových zrn v peří, jejich hustotou a rozmístěním v každém jednom peru na těle holuba. Na celkovém zbarvení a konečném výsledném dojmu se spolupodílí i lesk a svit, vznikající odrazem světla a jeho rozkladem ve stavebních strukturách jednotlivých per. Pro praktické využití dosavadních poznatků strohé genetické teorie související se zbarvením holubů je dobré výklad vést na příkladech známých a snadno pochopitelných. Každý holubář a chovatel je ve své podstatě šlechtitelem, který uplatňuje záměrné připáření a následně selekci, neboli výběr nejlepších jedinců jako rodičů další generace. Aby se genetici, šlechtitelé, ale i holubáři domluvili a věděli vždy jednoznačně o čem se baví, musí přijmout určité názvosloví a určitou odbornou terminologii.
Dominance a recesivita
Všichni z vlastní praxe víme, že některé projevy znaku převládají neboli dominují nad projevy znaku, které lze potlačit neboli překrýt. Chápeme většinou teorii, že vloha je dominantní neboli převládající, někdy lze doslova říci i překrývající účinek vlohy druhé, která je potlačitelná neboli ustupující – odborně řečeno vloha je recesivní. Další ne zcela často připomínanou skutečností je stav, kdy v heterozygotní konstituci může existovat neúplná dominance, kdy dominantní alela převládá svým projevem nad projevem alely recesivní, ale na fenotypu tohoto heterozygotního jedince poznáme, že se na konečném projevu znaku podílí svým účinkem obě vlohy z obou genů na chromozomovém páru a to jak alela čili vloha dominantní, která nezcela překryje současně se projevující účinek recesivní druhé alely.
S těmito pojmy se musí každý šlechtitel a tedy i holubář naučit pracovat a hlavně jim plně rozumět. V praxi si význam těchto pojmů vysvětlíme na zbarvení peří holubů. Ron Huntlay na svých webových stránkách http:// www.angelfire.com/ga/huntleyloft/ Page1.html, uvádí 38 různých genů, které jsou v odborné literatuře popsány a ovlivňují výsledné zbarvení peří holubů. V rámci některých genů jsou navíc pospány i vícečetné alelické série, tedy existence více než dvou odlišných alel v tomto jednom sledovaném genu.
Takovým genem je i gen „B“ určující barvu pigmentových zrn v peří holubů se třemi alelalmi. Recesivní neboli potlačitelná alela „b“ způsobuje hnědou barvu pigmentových zrn a určuje skupinu holubů hnědých a hnědožlutých. Tato alela „b“ se může uplatnit a projevit hnědým zbarvením peří jedině pokud bude u samců na obou pohlavních chromozomech ZZ. (obdoba chromozomů XX u samic savců). Recesivní alely se vždy zapisují malým písmenem (bb) a dominantní alely prvním velkým písmenem. (B+B+) nebo (BABA) v našem případě genu základního zbarvení pigmentových zrn. U holubic je vždy tato alela na jediném Z chromozomu, protože druhý pohlavní chromozom W tento gen nenese. (Obdoba Y chromozomu u samců savců genotypu XY) U holubů samců hovoříme v tomto případě dvou stejných alel o homozygotním stavu (bb) a u holubic jde vždy u genů na pohlavním Z chromozovu z pohledu pouze jedné alely o stav hemizygotní (b–)
Další alelou v tomto genu je dominantní vůči alele „b“ alela „B+“ vytvářející základní černou barvu pigmentových zrn. Dominantní projev znaku znamená, že černá pigmentová zrna budou nejen v případě homozygotních jedinců (B+B+), ale i v případě heterozygotních jedinců genotypu (B+b). Černá pigmentová zrna určují základní skupiny barev holubů černé-modré a šedočerné- stříbřité. Třetí a nejvýše z hlediska dominance stojící alelou v tomto genu je alela (BA). Výsledkem jejícho působení na jednom nebo i na obou chromozomech holubů samců je tvorba červených pigmentových zrn a to ve všech třech možných alternativách. V homozygotním stavu (BABA) a také v obou možných heterozygotních situacích (BAB+) a také (BAb). Červená pigmentová zrna určují skupiny popelavě červených a popelavě žlutých základních barevných skupin holubů.
Z výše uvedeného lze odvodit , že samci genotypu (BAb) budou ve svém potomstvu vždy dávat 50 % holubiček s červenými pigmentovými zrny (BA–) a 50 % holubiček se hnědými pigmentovými zrny (b–). Stejně tak budou štěpit na holubičky s červenými nebo černými pigmentovými zrny heterozygotní samci genotypu (BAB+).
Tito červeně popelaví heterozygotní holubi jsou i příkladem neúplné dominance alely „BA“ vůči alele „B+“, protože se v jejich červeně popelavém zbarvené objevují černé či modré skvrny jako projev překrývané alely „B+“, které se ale nevyskytují na homozygotních holubech genotypu (BABA).
S ohledem na rozsah článku nebudu veškerých 18 možných křížení holubů a holubic jednotlivých stejných nebo odlišných genotypů rozepisovat do tabulek a punnetových neboli mendelovských čtverců, ale každému zájemci o pochopení práce s alelami a geny to vřele s tužkou v ruce a s dostatečně velkým papírem doporučuji. Jednoduchá rada je do jednoho řádku si napsat alely zastoupené u holuba samce a za písmenko x jakožto krát vpravo si napíšete alelu u holubice. Na druhý řádek si napíšete možné genytypy a pohlaví potomstva. Například holub červeně popelavý heterozygotní (BAb) x holubice černá (B+ –) dají možné holuby popelavě červené heterozygoty (BAB+)a holuby černé heterozygoty (B+b) a dále holubice hemizygotní červeně popelavé (BA–) a hnědé (b–). Podobně se dá rozepsat, vysvětlit a dokázat genetický základ a výsledný barevný projev na rodičích a na jejich potomstvu. A právě zde je dobré se vrátit k připomenutí nezbytnosti vedení rodokmenů a s podrobnými zápisy zbarvení (ale i jiných vloh) u veškerého potomstva každého holuba i holubice na holubníku, pokud chceme svoji šlechtitelskou práci dělat zodpovědně.
Jak z obecné literatury a holubářské praxe všichni víme existuje u holuba další velice důležitý gen pro vytváření kreseb a vzorků na křídelních štítech všech základních barev holubů. Je to gen (C), který existuje minimálně ve čtyřech různých formách neboli vlohách čili alelách pro konkrétní projev účinků tohoto genu. Je to vloha „CT“ pro tmavý vzorek, vloha „C“ pro kapratost a dále vloha pro vzorek pruhový označená „C+“ a vloha „c“ pro bezpruhost. Na tomto příkladu genu se 4 známými vlohami si vysvětlíme možné vzájemné vztahy těchto vloh. Na nejvyšším stupni dominance v této alelické série stojí vloha „CT“ pro vzorek tmavý, která je vždy v pozici dominantní vůči všem ostatním alelám. Prakticky to znamená, že účinek této vlohy překrývá vždy účinek ostatních vloh stojících níže v této alelické sérii. Stejně tak vloha pro kapratý vzorek „C“ překrývá účinek vlohy „C+“ a vlohy „c“. Vloha pro pruhový vzorek „C+“ překrývá pouze účinek vlohy pro bezpruhost „c“.
A zde často začíná nechuť a odpor ke genetice, která se může v tomto momentu zdát příliš složitou a nejednoznačnou. Mnozí se nechtějí smířit s tím, že vloha například pro vzorek tvořící pruh zapisovaná „C+“ je dominantní vlohou ve vztahu ke vloze odpovědné za bezpruhý ráz „c“ a současně je vlohou v recesivním vztahu k vloze „C“, to je k vloze pro kapratý vzorek a stejně tak k vloze „CT“ určující tmavou kresbu či vzorek. Navíc výše uvedené popisy vycházejí z odborné literatury. Ve skutečnosti ale může nastat situace nepopisovaná v literatuře, kdy mohou některé projevy těchto vloh genu (C) být i neúplně dominantní a v detailním pohledu zkušený chovatele může vyčíst z fenotypu sytosti například kapratého vzorku odlišný genotyp (CC) oproti genotypu (CC+) nebo (Cc). Podobně zkušený chovatel praktik odhadne zda u pruhového holuba se jedná o homozygota (C+C+) se silnějším sytě probarveným pruhem oproti heterozygotovi (C+c) se slabší kresbou pruhem.
Pro naprosté pochopení práce s genetickými zápisy tak zvaných genetických vzorců, je třeba připomenout a znovu zopakovat, že každý holub i holubice mají na jednom daném příslušném chromozomovém páru vždy pouze dvě vlohy a to vždy jednu vlohu na každém jednom chromozomu, kterou zdědily od otce přes spermii a jednu vlohu na chromozumu od matky, který získali přes neoplozené vajíčko. Za genetický vzorec neboli za genetický zápis můžeme považovat i zápis dvou alel v jednom genu. V tomto příkladu může nastat deset možností genotypů, tedy alternativ genetického základu a celkem čtyři obecné fenotypy, to je vnější výsledný vzhledový projev účinku daného genotypu na jedinci.
Čtyři fenotypy a jejich možné genotypy jsou následující pro tento gen. Fenotyp vzorku tmavý může mít tyto genotypy (CTCT), (CTC), (CTC+) a (CTc). Homozygotní jedinec je genotypu (CTCT), neboli je nositelem stejné alely od obou rodičů. Ostatní jsou heterozygoty, neboli nositeli různých vloh od svých rodičů. Druhý fenotyp neboli výsledný vzhed je vzorek kapratý s možnými třemi genotypy = (CC) homozygot a dva heterozygoti (CC+) a (Cc). Třetí fenotyp pruhový vzorek má pouze dva možné genotypy (C+C+) homozygot a (C+c) heterozygot. Poslední možný čtvrtý fenotyp bezpruhý může existovat pouze v jediném hogozygotně recesivním stavu to je dle genetického zápisu do genetického vzorce (cc).
Malý příklad na závěr. Doporučuji každému si vzít tužku a papír a rozkreslit si sám pro pochopení a odvození příklad křížení heterozygotního holuba tmavého genotypu (CTC+) s holubicí kapratou heterozygotní s alelami (Cc). V potomstvu můžeme dostat potomstvo tří fenotypů neboli vzhledu a to „tmavé“ genotypů (CTC) a (CTc). Dále potomstvo „kapraté“ (CC+) a „pruhové“ (C+c). Pro bližší vysvětlení a využití tohoto příkladu i pro jiné modelové případy, ale i pro konkrétní chovatele i pro případné jejich konkrétní situace na holubníku, připomenu vlastní podstatu procesu rozmnožování, kdy výše v tomto odstavci uvedený holub genotypu (CTC+) vytváří spermie, které nesou buď vlohu „CT“ nebo vlohu „C+“. Jiné spermie tento holub nevytvoří. Holubice genotypu (Cc) v neoplozených vajíčcích před jejich oplozením má pouze buď vlohu „C“ nebo vlohu „c“. Proto tento pár bude dávat výše popsané čtyři možné genotypy a současně i 4 možné fenotypy potomstva.
Po prostudování těchto dvou příkladů vzájemných vztahů vloh neboli alel v rámci jednoho genu postoupíme ke studiu a hlavně k využití vzájemných účinků více genů. Postoupíme v genetickém vzorci zbarvení holubů dále od genu pro barvu pigmentových zrn (B) ke genu určujícímu rozmístění pigmentových zrn v peří a tím je gen (S) = (Spread anglicky), což by česky mohlo znamenat rozprostřený nebo překrývající, čímž je vysvětlen jeho účinek a vztah ke genu (C) základních vzorků. Alela recesivní „s“ v homozygotní konstituci (ss) nemá schopnost překrývání jako alela dominantní „S“ a díky tomu umožní, aby se v peří jedince genotypu (ss) uplatnil účinek různých alel genu C pro vzorky. Naproti tomu homozygotní konstituce (SS) plně překrývá účinek genu (C) a vytváří jednotné zbarvení bez vzorků a tím i celobarevný štít a letky křídla.
Doposud jsme výše hovořili o vzájemném vztahu dominance a recesivity dvou vloh jednoho genu. Nyní se začneme věnovat vzájemným vztahům a ovlivňování účinků více genů.
Vztah dvou genů hodnotí tak zvaná epistaze. Gen (S) je dominantně epistatický vůči genu (C) protože v dominantním zastoupení alely „S“ při genotypu (SS C?C?) překryje jakýkoli účinek jakéhokoli zastoupení alel v genu (C). Proto je za písmenem C otazník, který může vyjadřovat dvě situace. Jednak , že v genu C může být jakákoli alela, bez významu pro finální vzhled fenotypu a nebo že díky překrytí účinkem dominantní alely „S“ genu Spread netušíme, jaké alely v genu (C) na konkrétním holubovi jsou zastoupeny.
Opět si dovolím konstatovat, že výše uvedené je obecnou teorií. Ve skutečnosti může nastat rozdílný účinek překrytí, pokud plnobarevný holub bude nositelem homozygotní konstituce (SS) nebo pokud bude heterozygotem (Ss). Z vlastní zkušenosti znám situace, kdy zejména na výletkovi před přepeřením do peří dospělého jedince je na první pohled možné odhadnout, že se jedná o heterozygota a jemožné též odhadnout jaké alely jsou zastoupenu v genu C pro vzorek na štítu křídel.
Jak je zvykem ve skutečné vědě, bývá vědecky prokázána celá řada výjimek, potvrzujících jednotlivá vědecká pravidla. Tak například účinek dalších genů a vhodné konstituje jejich některých alel s názvem Toy Stencil v konstituci (Ts1Ts1, Ts2Ts2, ts3ts3) způsobí možný projev převládnutí bílé kresby vzorků řízených genem C nad účinkem genu S (Spread). A to i v jeho homozygotní konstituci (SS) a umožní prosazení se vrozků v bílé formě na plnobarevném štítu při zachování plnobarevné letky. U moravského pštrosa v současnosti je to kresba černá bělopruhá. Ta vznikla přenesením těchto vloh genu Toy Stencil z jiných plemen, nejspíše z Polského rysa, tělesným rámcem nejbližšího Moravskému pštrosu. Tato forma bělopruhosti by se dala přenést i do ostatních barev modré, červené a žluté, případně hnědé i stříbřité. Jednalo by se o jiný genetický podklad a také o jiný fenotypový projev bělopruhosti oproti současnému stavu, kdy u bělopruhých rázů moravského pštrosa se uplatnujě alela
„Od“ genu Dominantní Opál, který pouze vyběluje tmavou část vzorků u vlohy pruhové (C+) nebo kapraté (C) u kresby šupkaté a navíc zesvětluje letky u červených a žlutých a u modrých vyběluje černé části pigmentace letek, někdy až do barvy zcela bílé. V případě uplatnění alel komplexu Toy Stencil by to znamenalo mít bělopruhého modrého nebo i žlutého moravského pštrosa s tmavou letkou stejnou jaká je u plnobarevného rázu, na rozdíl od stávajících probělených letek při uplatnění alely „Od“. Holubi bělopruzí u moravského pštrosa by tak mohli tvořit dva geneticky odlišné kresebné rázy. Bělopruhost „typu Ts“ by ale byla mnohem náročnější na zušlechťování díky nutnosti vhodného uspořádání alel v komplexu tří genů, který se naruší při nezbytné potřebě křížení se špičkovými jedinci základní bezpruhé barvy a proto jejich šlechtění je velice náročné a získání dokonalého jedince je skutečným mistrovkým dílem. Je nutné si uvědomit, že jedna ze tří potřebných konstitucí alel v těchto „Ts genech“ je recesivní v genu (ts3ts3), kdy pouze toto recesivní zastoupení alel umožní probělení vzorků do té doby skrytých pod účinkem genu Spread (konstituce SS). Odborně řečeno se nám tady uplatňuje recesivní epistaze.
Na těchto dvou příkladech účinků více alel jednoho genu a příkladů vzájemných účinků různých alel více genů nechci dokumentovat obecně složitost genetiky, naopak chci poukázat na potřebu důkladné přísnosti v selekci a potřebu dokonalé evidence projevu znaků u všech potomků v každém vlastním chovném páru v průběhu celé sezóny. Tyto vlastní poznatky je vhodné potom využívat jako podklad pro příští připařování právě díky znalosti, jaké potomstvo daný holub či holubice v předešlých sezonách dávali a s jakými protějšky.
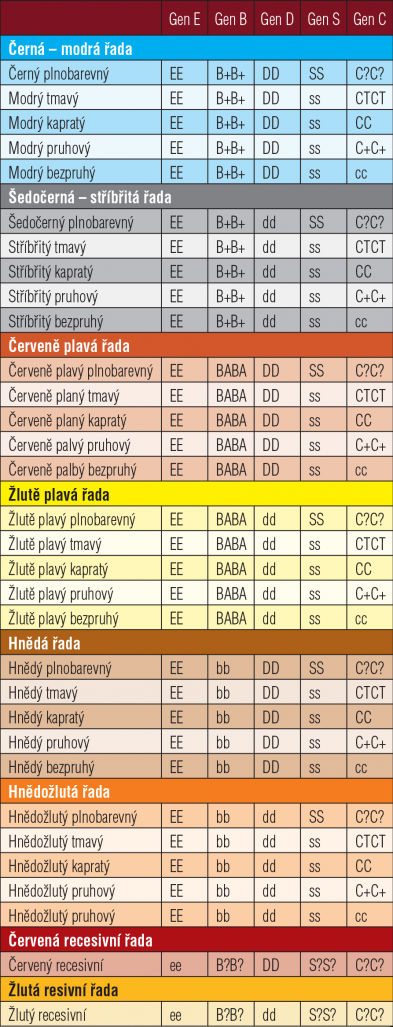
Pracovně je třeba z odborné holubářské literatury přijmout existenci genetického vzorce pro základní zbarvení a kresby u holubů zhruba v podobě uvedené v následující tabulce č. 1. Omezíme se v ní pouze na geny pro barvu peří a případný vzorek. Otazník za zápisem genu vyjadřuje možnost zastoupení jakékoli alely v daném genu v případě dominantně epistatického účinku předcházejícího genu. Jen ve stručnosti se ještě zmíníme o dvou genech uvedených v Tabulce č. 1. Gen (E) pro recesivní červenou nebo žlutou barvu způsobuje červené zbarvení při recesivní epistazi to znamená při homozygotním zastoupení obou alel v genotypu(ee). Tento epistatický účinek způsobí červené zbarvení peří bez ohledu na zastoupení alel v genu pro zbarvení pigmentu (B). Jeho vzájemný vztah k dalším genům je opět ovlivňován současným spolupůsobením mnoha dalších genů a je nad rámec tohoto článku zabíhat do podrobností. Tento gen (E) recesivní červené a žluté barvy zmiňujeme pouze pro jeho podíl na jednom ze základních typů zbarvení.
Poslední gen zatím nezmíněný, ale velice významný pro základní zbarvení holubů je gen (D) dilute z anglického termínu, znamenající rozředění, který ovlivňuje hustotu pigmentu v peří a z intenzivních barev – černé (modré), popelavě červené, hnědé a recesivní červené při recesivní konfiguraci (dd) u holubů samců a (d–) u holubice vytváří zbarvení základní rozředěné – šedočerné (stříbřité), popelavě žluté, hnědožluté a recesivní žluté.
Zastoupení vloh v genech řídících základní zbarvení holubů
(Pokračování příště)
RNDr. MILAN TYLLER,
vzdělavatel KCHHMP Brno